人类细胞老化的速度和自由基攻击细胞後产生的代谢物有明显相关,而能够清除自由基力的SOD则成为健康水平和寿命长短的指标。研究显示,灵芝萃取物不论在体内外都有减少自由基、提升SOD活性的能力;灵芝本身甚至含有大量的SOD组成蛋白质,可以作为SOD的补充来源,令其延缓衰老的机制更加全面。
文.图/许瑞祥
◎本文原载於1999年《健康灵芝》3期19~22页
人类的衰老过程至今仍是一个谜,因此无法任意控制。多少世纪以来,方术之士努力不懈找寻长生不老的药方,虽然都没有成功,但在各国的历史中却留存着人类追求青春永驻的深刻记忆。传言中的秦始皇为求长生不老,派出大批的童男童女远赴海外寻访仙草,成为中国历代皇帝寻求不死灵丹的滥觞。
根据正史记载,宋真宗赵恒是第一位服食灵芝以期能长生不老的皇帝,而宋徽宗赵佶在位期间,则是搜刮灵芝的全盛时期。灵芝除了是皇宫大内的御用贡品外,中国古代的道士更将灵芝视为修练成仙的基本配备。在道教全盛的魏晋时期所编撰的灵芝专着不下百余种,由此可知灵芝在当时的重要性。
明朝李时珍综合历代本草的记载与道家的经验,在其所着的《本草纲目》中,对六芝有完整的叙述,无论是青、赤、黄、白、黑、紫中任一种灵芝,皆谓其能「久服轻身不老,延年神仙」,为自古以来灵芝抗老防衰的作用,作了最完整的阐述。
衰老和自由基有关
现代科学的研究认为,衰老的过程可能从生命诞生之时就已经开始,并且是一连串的不可逆反应。虽然减缓衰老的理论众说纷纭,但控制或清除体内自由基(free radical)的生成,却是目前医药学界共同努力的方向。
自由基是一个外层轨域含有一或多个不成对电子的原子或原子团所构成非常不稳定的状态。在含氧的生物体中,常见的是以氧分子为中心的超氧自由基(O2-)及氢氧自由基(OH.)。在真核生物的粒线体中,因不正常的电子传递结果,将一个电子交给氧分子而形成O2-【1】。
根据Turrens等人对牛的心脏细胞分析结果显示,其中的粒线体在进行电子传递链之作用时,会有3~5%的氧形成超氧自由基(O2-)【2】。
曾有人预估人类每天呼吸的氧气中约有1~3%变成O2-,而此类不稳定的自由基生成後,常会攻击其周围正常的分子,产生连锁反应,对生物细胞造成许多的伤害,包括引起细胞膜中不饱和脂肪酸的过氧化,继而改变细胞膜的通透性;或因加速细胞内DNA的氧化伤害,导致基因突变机率的增加等,皆与细胞突变、老化或死亡有密切的关系。最新的研究显示,人类细胞老化的速度和自由基攻击细胞後产生的代谢物有明显的相关性【3】。
SOD是体内清除自由基的主要酵素之一
为了对抗自由基对细胞所造成的伤害,好氧性生物体的正常细胞内外都有防御系统。细胞内负责消除超氧自由基的酵素系统包括:Superoxide dismutase(SOD)、Glutathione Peroxidase和Catalase三种酵素。
另外非酵素系统中的抗氧化剂,如维他命C(vit C)、维他命E(vit E)等,亦可直接与自由基作用,进而防止连锁反应的发生。在好气性生物细胞中的超氧歧化酶(SOD)於1969年被McCord和Fridovich发现,它是一群含有金属元素的酵素,负责清除超氧自由基的反应如下【4】:
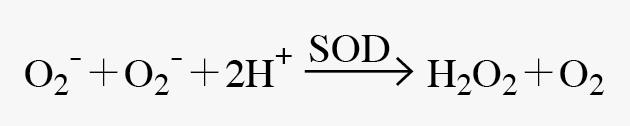
目前已知的超氧歧化酶(SOD)依其所含金属离子之不同可分为三大类:
1. Cu/Zn-SOD:主要存在细胞质和叶绿体中。
2. Fe-SOD:主要存在原核生物或植物叶绿体中。
3. Mn-SOD:主要存在真核生物的粒线体中。
监於超氧歧化酶(SOD)为好气性生物细胞对抗过氧化毒害的第一线防卫系统,因此SOD的活性高低成为评估生物与环境互动的指标。在植物的研究中发现,当绿色植物突然受到强光照射时,光合作用的速率瞬间提升,使得在光反应系统中部分电子未能顺利交给NADP,而由O2接受形成超氧自由基O2-。
但当植物遭受环境剧变时,其体内测得的SOD活性有增加的趋势,因此植物体内SOD活性的变化,常成为评估植物遭逢逆境时的应变能力【5】。
在老鼠皮肤细胞的评估模式中发现,细胞中的SOD活性会随着细胞年龄的增长而逐渐降低。1990年Niwa等人所提出的临床测量分析结果证实,人体内的SOD浓度较低或其SOD活性较差者,通常其寿命较短或罹患疾病的机率较高【6】。从此SOD不但成为人类健康等级与寿命长短的客观指标,同时也成为抗老防衰的灵丹妙药,各种动、植物来源的SOD产品亦成为现代抗氧化、防衰老的新宠。
本文将以清除超氧自由基的超氧歧化酶SOD为主要探讨对象,以现代抗衰老的药理研究基础,来重新检视久食轻身不老的灵芝,是否具备了符合潮流、令人确信的能力。
灵芝萃取物能提升SOD活性
根据上海医科大学药学院所发表的试验结果,灵芝的粗萃取物在试管中能直接清除超氧自由基(O2-),且其作用呈现剂量的相关性。由此可知,在灵芝的萃取物中含有类似抗氧化剂的成分。在动物体内试验中,经灵芝萃取物灌胃的大白鼠,分析其血液中红血球的SOD活性,结果显示以500 mg/kg、250 mg/kg、125 mg/kg灵芝萃取物灌胃後的各组,皆呈现SOD活性提高的现象,同时发现其红血球细胞膜的流动性与封闭度,较未经灌食灵芝萃取物的各组皆有明显的提升【7】。
在已知疾病形成的过程中,常会见到细胞膜流动性发生改变。通常红血球细胞膜的流动性亦会随着细胞年龄的增长而降低,因此能提高细胞膜的流动性有延迟细胞衰老的意义。红血球的封闭能力也与细胞年龄有关,年轻的红血球其封闭度为93%,但老化的红血球其封闭度仅为36%【8】。由此可知,若能提高老化红血球的封闭度,对细胞抗老化作用有积极贡献。
从前述喂食灵芝的大白鼠血液分析结果,显示在125~500 mg/kg剂量下,灵芝萃取物能明显提升红血球中SOD的活性,减少超氧自由基所造成的脂质过氧化反应。其作用的具体表现在血液分析中的平均粘度降低,以及细胞膜的流动性与封闭度的提高。
临床证据显示,以灵芝制剂治疗33例高血脂、高血压等病人两周後的血液流变学观察结果,病人的全血和血浆粘度明显下降,血液流变性改善【9】,对於提高心脑血液循环、预防血栓生成、降低血管系统疾病发生或发展有一定的临床意义。
灵芝含有大量的SOD组成蛋白质
灵芝自古以来一直是抗老防衰的代表,我们亦曾对灵芝菌丝体中的SOD活性进行系列之分析。在从两百多株食用真菌的菌丝体中筛选比较其中SOD活性的结果发现,灵芝属菌株皆具有很高的SOD活性,无论其种类或生长时期的改变,皆显示灵芝的菌丝体本身就是一种丰富的SOD来源。
从市场需求的角度,灵芝将可成为除了动物、植物来源的SOD外,另一个具有开发潜力的生物资源。服用灵芝不但可以刺激机体提升SOD的活性,同时灵芝本身亦含有大量的SOD组成蛋白质,灵芝延迟衰老作用机制的全面性,以现代科学的立场观之,更显得弥足珍贵。
自1994年起,我们开始进行灵芝属菌株中SOD基因与蛋白质组成构造的探讨,目前已经完成包括灵芝(Ganoderma lucidum)、松杉灵芝(G. tsugae)和小孢子灵芝(G. microsporum)等菌株的大量培养生产SOD的条件,并且深入探讨灵芝来源的SOD萃取、回收与纯化的过程【10】。
在已知的结果中显示,灵芝菌丝中的SOD是以Mn-SOD为主,纯化後的Mn-SOD在经70℃加热或以蛋白酶水解後,仍能保留其部分活性【11】。就利用开发的需要而言,灵芝来源的Mn-SOD确实是一种相当理想的SOD型式。
在有关其SOD基因的研究中,我们利用聚合酶连锁反应(PCR)来扩增灵芝属菌株的SOD基因片段。在1996年发表的小孢子灵芝(G. microsporum)Mn-SOD基因序列在当时成为人类基因资料库中,第一笔高等担子菌的SOD基因核酸序列。在分析比对30株灵芝属菌株中Mn-SOD基因资料时,发现其中皆含有两段非转译的插入序列(intron)。此两段插入序列的核酸定序结果经电脑进行演化过程的分析後,竟然得到与灵芝属菌株类缘关系一致的结果【12】。
从分子生物水平的分析确认在灵芝属菌株间特殊的SOD基因应该是来自於相同的起源,而每一个灵芝菌株的演化过程皆记录在此段Mn-SOD基因中非转译的插入序列中。
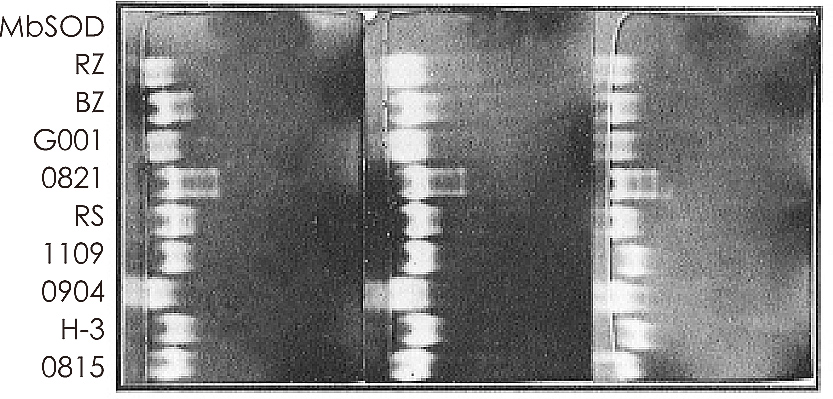
图(一) 灵芝属菌株超氧歧化酶(SOD)的同功酵素图谱。
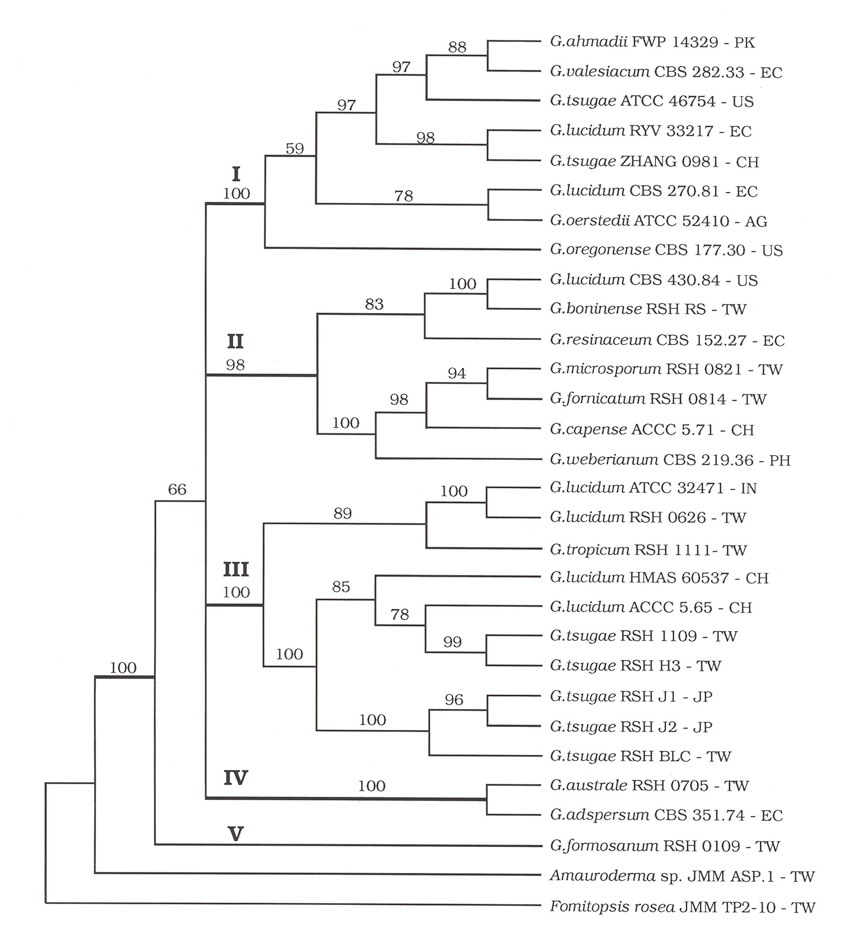
图(二) 利用Mn-SOD基因序列进行灵芝属菌株类源关系监定之结果。

图(三) 灵芝来源的Mn-SOD N端胺基酸序列特徵。
灵芝「轻身不老」有其科学依据
自1970年代开始进行灵芝抗衰老机制的系统化研究中,就自由基清除能力的评估模式来了解时,灵芝的萃取物在体外的试管中能直接消除其中的超氧自由基,而且有明显的抗氧化能力。经喂食後的寄主体内SOD活性提升,并且改善衰老红血球细胞膜的流动性与封闭度,使得几百年来口耳相传的经验,在现代严谨的评估下,重新得以确认。
另外,从蛋白质构造与基因相似性的证据,皆显示灵芝家族的成员其SOD组成的独特性。当我们了解灵芝延缓衰老的能力时,久食轻身不老将不再是遥不可及的神话。我们期待有更多的资源投入灵芝老年保健用品的开发,为银发族延续生命的第二个春天。
延伸阅读
积极开发灵芝研究,「延年益寿」将不再是神话
参考文献
1. Southorn, P.A. and G. Powis. 1998. Free radicals in medicine. I. Chemical nature and biologic reactions. Mayo. Clin. Proc. 63 : 381-389.
2. Turrens, J. F. and A. Boveris. 1980. Generation of superoxide anion by the NADH dehydrogenase of bovine heart mitochondria. Biochem. J. 191 : 421-427.
3. Halliwell, B. Antioxidants. 1994. Sense of speculation? Nutrition Today. 29 : 15-19.
4. Mc Cord., J.M., and I. Fridovich. 1969. Superoxide dismutase. An enxymatic function for erythrocuprein. J. Biol. Chem. 244:6049-6055.
5. Karpinski, S., et al. 1992. Differential expression of Cuzn-superoxide dismutase in Dinus sylvertris needles exposed to Soa No2. Physiologia plantarum 85 : 689-696.
6. Niwa, Y., et al. 1990. Induction of superoxide dismutase in leulsocyter by paraguat;correlation with age and possible predicton of longevity. Blood 76 (4) : 835-841.
7. 李端等,1994,灵芝对大鼠红细胞超氧歧化酶(SOD)和膜蛋白组成的影响。灵芝的研究(一),pp.92-95。
8. 李端等,1994,灵芝对大鼠细胞膜流动性和封闭度的影响。灵芝的研究(一),pp.86-91。
9. 程彰华等,1994,灵芝对高血压伴高血脂和脑血栓後遗症患者血液流变学的观察。灵芝的研究(一),pp.100-103。
10. 叶志新,1996,灵芝属含锰超氧歧化酵素的纯化与定性。国立台湾大学农业化学研究所硕士论文,p.123。
11. Pan. S. M., et al. 1997. Purification and Characteriza of Manganese superoxide dismutase from Ganoderma microsporum. Biochemistry and Molecular Biology International 42(5):1035-1043.
12. 王惠芳,1996,灵芝属含锰超氧歧化酶基因之研究。国立台湾大学农业化学研究所硕士论文,p.107。

