历经1971至2005年共三十几年研究,科学家不仅从过去模糊的「高分子多糖体」,逐步厘清到「活性多糖肽」的概念,更找到这类活性成分调控免疫的直接证据。
文/许瑞祥
◎ 本文原载於2006年1月《健康灵芝》第31期26~29页
阳明大学许先业教授所发表的论文中证实,从灵芝子实体纯化而得
含有岩藻糖(9.7%)和15.6%蛋白质的活性多醣,可经由TLR4受体活化巨噬细胞。
(J. Immunol. 2004 Nov 15;173(10):5989-99.)
1997~2002, 韩国
1997 年韩国 Wonkwang 大学 Park 等人发表以灵芝多糖体抗肝纤维化的研究结果,发现在小鼠体内利用胆管结紮诱导肝纤维化的试验模式下,灵芝多糖可有效降低血清中 aspartate transaminase(AST,即 GOT)、alanine transaminase(ALT,即 GPT)、alanine phosphatase(ALP)和胆红素值,并可减低胶原蛋白在肝脏中的含量,此结果显示灵芝多糖体用於保肝药物的开发是具有潜力的。
1999 年韩国原子能研究所 Kim 等人发表灵芝(G. lucidum)子实体热水萃取的多糖体,具有抗超氧自由基伤害之能力,在体外的试管中对 DNA 有良好的辐射伤害保护能力,以及对抗氢氧自由基所诱发 DNA 断裂的能力。结果显示,灵芝可以为人类预防因自由基的攻击而引发的相关疾病,无论是癌症或是老化的发生,均能提供有效的预防途径。
同年,韩国 Chungbuk 大学 Eo 等人发表灵芝多糖体抗疱疹病毒的系列研究,结果显示灵芝子实体经热水萃取而得的多糖体中含有不同蛋白质键结的组成分,对於抗疱疹病毒的活性亦有差别。
在体外试验中,以溶菌斑减少法评估其对抗疱疹病毒Ⅰ型(HSV-1)和Ⅱ型(HSV-2)的活性,其中以含酸性蛋白质的多糖体具有最强的抗病毒能力,在 Vero 和 HE1-2 细胞株中 EC50 值为 520 μg/ml,作者认为具有开发成新抗疱疹药物的价值。
2002 年,同样的研究团队发表以此酸性蛋白键结的多糖体(APBP)搭配核苷型的抗疱疹药物Acyclovir(ACV)和 Vidarabine(ara-A)组合使用的抗病毒效果,利用 Combination Index(CI)值来评估多种药物共同使用的效果,结果显示 APBP 和 ACV 的 CI 值为 0.47~0.51,而 APBP 和 ara-A 的 CI 值则为 1.02~1.18。
此结果显示,APBP 和 ACV 具有明显抗 HSV-1 和 HSV-2 的协同作用,以此灵芝多糖搭配抗疱疹药物使用,不但可以减少使用剂量,亦可以提升其抗病毒效果,为灵芝搭配现有药物的使用提供有利的佐证。
同年,该团队的 Kim 等发表以此 APBP 搭配蛋白质型的抗病毒剂干扰素-α(Interferon-α, INF-α)和干扰素-γ(INF-γ)的效果,以 CI 值来评估不同组合抗疱疹病毒的结果,其中APBP 和 INF-α 对 HSV-1 和 HSV-2 的 CI 值为 0.3 和 0.62,APBP 和 INF-γ 对 HSV-1 和 HSV-2 的 CI 值为 0.65 和 1.10。
此结果显示,APBP 和 INF-α 的协同抗病毒作用较为明显,为灵芝多糖搭配干扰素使用於病毒的防治提供有效的证据,同时对於使用干扰素治疗的过程中,配合灵芝的共同作用,将可使干扰素的效果更为明显。
2002 年 Eo 等发表 APBP 抗疱疹病毒的作用机制,发现 APBP 在 100 和 90 μg/ml 时可以抑制 50% HSV-1 和 HSV-2 病毒附着於 Vero Cells 细胞上,同时可以阻止病毒渗透到Vero Cells 中。此结果显示,APBP 的抗病毒机制是经由结合细胞表面的 HSV 专一性附着的糖蛋白位置,阻碍了病毒和细胞接触的管道後,降低病毒入侵细胞膜的机率。
由灵芝多糖体抗人类疱疹病毒的系列研究成果,可见灵芝的热水萃取物能阻止病毒和细胞的专一性结合,减低细胞感染病毒的机会,同时在受到病毒感染的细胞以抗病毒药物治疗时,亦能与其发挥协同作用,提升药物的效果。此结果对於灵芝多糖体应用价值的确认,无论是预防或是治疗产品的开发,都有不可替代的重要性。
2002~2005, 台湾
2002 年中研院由翁启惠院士主导的灵芝研究团队,开始发表有关灵芝多糖体的系列研究成果。灵芝(G. lucidum)子实体经 0.1N NaOH 溶液萃取之上清液经中和後以乙醇沉淀的粗萃取物,其组成分析结果以葡萄糖(58%)和甘露糖(15.5%)为主,并含有岩藻糖(9.7%)、半乳糖(9.3%)和木糖(5.4%)等醣类外,并含有约 15.6% 的蛋白质。此粗萃取物再经以胶体滤管柱层析後,回收得到 5 个主要含醣的部分。
以小鼠脾细胞增生试验评估其主要作用的活性表现在第三部分(F3),在有效浓度为 0.01~1.1 μg/ml 时,对脾细胞有明显的增生作用,而原粗萃取物则需 0.1~1μg/ml 方可达到相同的效果。
以二维电泳分析小鼠脾细胞经 F3 处理後的蛋白质体变化时发现,约有 50% 的蛋白质产生变化。F3 促进脾细胞的增生外,同时亦可增加 IL-1、IL-2 和 INF-γ 的表现。作者强调在此多糖组成中的岩藻糖,对於活性的存在是必要的。
此篇论文显示,国内研究已经从基因体学进入到蛋白质体学的领域,对於活性多糖体的组成与构形具备精确描述的能力,同时显示此活性多糖是与蛋白质相结合的聚合物。
2004 年该团队中的 Chien 等发表以 F3(10~100 μg/ml)处理人类脐带血 7 天後,分析其中巨噬细胞、树突细胞和自然杀手细胞的细胞数目後,发现比未经 F3 处理的对照组分别增加 2.9、2.3 和 1.5 倍。此外,B 细胞数目没有明显的差别,但 T 细胞在 F3 处理下降低 20%左右,经 F3 处理後的自然杀手细胞,其毒杀癌细胞的能力增强 31.7%。
由此体外分析结果,确定含蛋白质的灵芝多糖F3不会伤害人类的免疫细胞,却可以促进细胞的增殖与提升其毒杀癌细胞的能力。另外,在本篇中首度提到灵芝多糖体能刺激人类脐带血中免疫细胞增殖的作用,提升来源与保存不易的脐带血在将来应用的可能性。
同年,该团队的 Chen 等发表将此具有活性的多糖体 F3,再经阴离子交换管柱层析详细分为F3G1(11%)、F3G2(10%)和 F3G3(11%)三部分,期待能找寻更小分子量的活性多糖体。
经以对小鼠脾细胞诱导生成细胞激素的模式来评估其活性的表现,结果发现 F3 能活化 IL-1、IL-6、IL-12、INF-γ、TNF-α、GM-CSF、G-CSF 和 M-CSF 等多种细胞激素。其中的 IL-1、IL-6 和 INF-α(前述不含此激素)与发炎反应有关,INF-γ、TNF-α 与抗肿瘤活性有关,GM-CSF、G-CSF 和 M-CSF 则与造血机能有关。
将 F3 再往下细分所得的各组分析结果:F3G1 只能活化 IL-1、IL-12、TNF-α 和 G-CSF 四种细胞激素,F3G2 和 F3 相同能够活化前述8种细胞激素,F3G3 则只能活化 IL-1 和 TNF-α 两种,结果显示 F3G2 和 F3 有相同的作用机制与活性,但 F3G2 的分子量明显较 F3 为低。
作者等在论文中虽未曾提及有关 F3G2 和 F3 在组成与构形分子量间的差别,但就此结果而言,具有启动免疫调节能力的多糖体,其基本结构已被精确掌握,从灵芝的粗萃取物中可精制纯化得到多少量的 F3G2 是可以计算的。
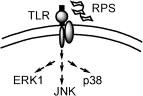
同年该研究团队并确认,灵芝多糖体启动免疫调节的关键,是经由细胞表面特定受体来达成的。阳明大学许先业教授所发表的论文中证实,F3 是经由与巨噬细胞表面 TLR4 受体的结合来启动细胞内 IL-1 基因的表现,不但排除其他来源的 LPS 干扰,亦推翻过去认为是经由Complement receptor type 3 启动的说法。
无论是人类或小鼠的巨噬细胞,F3 皆可经由细胞表面上的 TLR4 受体,向细胞内调控 IL-1 表现相关的基因与酵素。在细胞分子层级的分析结果中可见,当 F3 与 TLR4 受体结合後,立刻活化细胞内的 PKC、MEK1、PAK、MAPKs 等酵素系统,将不同的讯息向细胞核传递,进而引发多种细胞激素的生成。
本文中以 IL-1 的表现与调控为指标,建立评估活化或启动路径的网路团,对於 IL-1 基因於表现前转录与 F3 调控後转录的 pro-IL-1,到 IL-1 的生成与分泌至胞外的过程,皆有清楚的了解,对於灵芝多糖体於细胞外如何经由特定 TLR4 受体来启动胞内的基因表现,生成细胞激素後分泌到胞外参与免疫调节的过程,有更精确的认识。
大约在同时期,另一组台大医学院和阳明大学合作的团队亦发表系列多糖体的研究成果。2002 年 Hsu 等发表以灵芝多糖体延长白血球的存活时间来增强免疫能力的论文,由灵芝(G. lucidum)子实体经热水萃取、酒精沉淀的粗多糖,再经过胶体管柱层析与阴离子交换树脂管柱分离纯化所得的高分子多糖体代号为 PS-G,经详细的组成分析确认 PS-G 应该是由 95% 的多糖体和 5% 的胜肽所结合而成的多糖肽,同时已知此 PS-G 不含细菌内毒素,以排除受 LPS 污染的干扰。
作者等研究 PS-G 对嗜中性白血球存活率的影响,是透过抑制自发性凋亡现象达到的。嗜中性白血球是重要的免疫细胞之一,主导寄主抗菌与吞噬的能力,此白血球从骨髓释放到循环系统中只有 6~10 小时的生命期,受到自发性凋亡程序来调控其生命的长短。研究发现,PS-G 能拮抗由 Akt-regulated signaling pathways 所引发的白血球凋亡作用,延长白血球存活时间来增强寄主免疫力。
2005 年 Lin 等发表以此多糖肽 PS-G 启动人类树突细胞的机制,证实 PS-G 亦是经由树突细胞表面的 TLR-4 受体将讯息传入细胞内,使负责讯息的蛋白分子 NF-KB 和 p38MAP 等传递途径加速树突细胞成熟,同时合成并分泌细胞激素 IL-12。IL-12 不仅能增强自然杀细胞和 T 细胞噬菌的能力,促使自然杀手细胞分泌 INF-γ 外,还能诱导 T 细胞朝向 Th1(第一型免疫反应)分化。
灵芝多糖体启动免疫讯息的起始点,在人类树突细胞的研究模式中再次获得证实,就作者的描述 PS-G 和 F3 都是含有蛋白质的多糖体,其来源不同,分离纯化条件各异,虽然皆未标示活性多糖体的分子量大小,但就无论是巨噬细胞或是树突细胞,皆以 TLR-4 受体结合後开始启动免疫讯息的结果而言,此两种活性多糖肽的基本构造必定是相似的。
2002~2005, 中国
中国北京大学药学系林志彬教授组织的研究团队,在此时期有关灵芝多糖体的研究亦大放异彩。在其系列相关研究所用的材料是以灵芝(G. lucidum)子实体经热水提取、酒精沉淀、透析与去蛋白质等程序後所得的Gl-PS(G. lucidum polysaccharide),对应於子实体乾重的产率为 0.82%。
此多糖经精密的组成分析後,确认应该是多糖肽(polysaccharide peptide),其分子量为 584,900,多糖和肽的比例为 93.51:6.49。其中多糖由鼠李糖(rhamnose)、木糖(xylose)、果糖(fructose)、半乳糖(galactose)、甘露糖(mannose)和葡萄糖以 0.793:0.964:2.944:0.167:0.384:7.94 比例组成,多肽的部分则由17种胺基酸所组成。
2002 年该团队 Cao 等发表以 Gl-PS 处理树突细胞後,不但能加速其成熟,同时可以诱导毒杀性 T 细胞(Cytotoxic T-lymphocytes, CTL)细胞毒性的生成,是经由 INF-γ 和 Granzyme B pathways 来调控。
2003 年 Zhang 等发表 Gl-PS 无论在体内或体外试验结果,显示 Gl-PS 可以保护胰岛 β cells 不受自由基的伤害,为多糖体抗自由基活性提供另一项有利的佐证。
2004 年 Shao 等针对灵芝多糖体启动免疫反应的受体位置发表其研究成果,以 Gl-PS 处理能活化 BALB/c 小鼠的 B 细胞和巨噬细胞,但不能活化 T 细胞;TLR-4 受体突变的 C3H/HeJ 小鼠的 B 细胞则无法被 Gl-PS 活化。
作者发现利用抗小鼠 TLR-4 的单株抗体可以有效抑制被 Gl-PS 刺激後 BALB/c 小鼠 B 细胞的增殖现象。综合以上结果可知,TLR-4 受体是 Gl-PS 活化 B 细胞的必要媒介。
在巨噬细胞的试验结果,Gl-PS 可以诱导 BALB/c 小鼠腹腔巨噬细胞产生 IL-1β,但对於 TLR-4 受体缺损的 C3H/HeJ 小鼠则无法产生诱导作用,显示 Gl-PS 活化巨噬细胞时需要有 TLR-4 受体的参与。
本论文以细胞表面具有 TLR-4 受体与不具 TLR-4 受体的 B 细胞和巨噬细胞互相交互验证,灵芝多糖体要调控免疫,确实是经由 TLR-4 受体结合後向胞内传递讯息。
总结
自 1970 年开始有关灵芝活性成分的研究报告中,多糖体一直是被探讨的主角,从开始的抗肿瘤(S-180)活性到提升免疫功能,1990 年从细胞激素的消长了解灵芝多糖体具备免疫调节功能。
时至今日,对於启动免疫反应所需的特定受体与讯息传递途径已有明确的结果,无论是树突细胞、巨噬细胞或是 B 细胞,被活性多糖肽 Gl-PS、PS-G、F3 等启动的皆为细胞表面的 TLR-4受体,为高分子量的多糖参与免疫调控找到直接的证据,同时也让我们从过去模糊的高分子多糖体,逐步厘清到活性多糖肽的概念。
确实有效多糖肽的基本构造或许仍然未知,或许是重要的商业机密,但对大部分的读者而言,了解灵芝功效中抗癌、抗氧化、抗病毒、降血糖、护肝与免疫调节等,皆来自於特定组成与构形的多糖肽。
当我们受益於不断提升品质的灵芝产品时,更应该对三十多年来埋首研究灵芝多糖体的各国精英表达无限的敬意与谢忱。由於篇幅有限,本人谨对许多尚未被引用的遗珠之憾表达歉意。

