多醣是灵芝,乃至其他食用药用真菌的主要成分。就菇本身而言,为什麽要形成多醣?其化学结构有什麽特性?有哪些生理功能?台北医学大学教授苏庆华不仅回顾了他在1995~2000年利用菌丝发酵生产灵芝多醣提供董大成院长做抗肿瘤实验的有趣经历,也回顾了他对於子实体中水可溶、硷可溶与强硷可溶多醣的探索。在这一步步挖掘的过程中,他最大的感想是,灵芝真是好东西,每一个萃取阶段都有好的成分展现出来,而最好的多醣居然留在最後才出现……
文/苏庆华
◎本文原载於2014年《健康灵芝》第63期 4~6页
从1980年代开始,真菌代谢的多醣(polysaccharides)开始受到日本学者的关注,例如最早被拿来做实验的香菇多醣(lentinan)、云芝多醣(krestin),这两种多醣的抗肿瘤研究逐渐为学界接受,并开发成为临床用药。
因此之故,许多安全上已被认可的食药用真菌所代谢之多醣,也陆续被拿来做比较,一系列描述木耳多醣、白木耳多醣、裂摺菌多醣(schizophyllan)、竹荪多醣……等等的论文相继问世。灵芝身为药用真菌之要角,自然无法置身事外,最早有关灵芝多醣的文献即在此时出现了(Mizuno et al., 1985)。
建立多醣分子属性的分析方法
多醣是真菌细胞壁主体的成分,也是构成真菌细胞壁的主要成员。多醣与几丁质(chitin)互相交错堆叠,形成一个具有弹性并赋予真菌细胞形状的外壳,不仅能保护细胞,还具有通透性,使真菌细胞可以透过它来吸收水分及小分子养分,甚至能阻止具有破坏性的大分子(如分解性酵素, lytic enzymes)入侵。
虽然真菌细胞壁所构成的多醣有这些共通性,但不同种真菌所产生的多醣极其复杂,这也是研究上需要花许多力气的原因。首先,多醣的合成过程不像蛋白质有一定的氨基酸序列及大小,同样一组多醣合成酵素,受到养分供应及环境因素的影响,会多加或减少单醣接到多醣之长短;其次,这些酵素也会因为特异性不同而接上不同的单醣;连接时,架键的位置也会改变。
为此,科学家们设计了一套方法来描述多醣的分子属性。第一个当然就是分子大小,常用的方法就是把多醣用固定大小的分子筛过滤或层析,再用已知大小的标准品做回归求得。
有了已知大小的多醣後,取其中一部分用酸分解成单醣,检测是哪一种或哪几种单醣,其比率如何;然後取另一部分多醣,测定醣与醣之间是怎麽结合的,是1→2、1→3、1→4或1→6;最後再用酵素确定这些醣结合的方式是α-还是β-。
有了这些资料,我们就可以知道这个多醣的基本结构,例如β-(1→3)d-glucan即代表「具有由右旋(d)β葡萄糖、以1→3键构成之多醣」。
当初为了研究多醣,我花好几年的时间才把仪器设备及特别的试药程序建立起来。那时我刚到北医,没有玩过多醣,在大家都很热中多醣的大环境下,尝试自我学习。但不久之後,灵芝多醣已经被研究过了,在学术上失去了新颖性,所以当我开始研究多醣时,就锁定了另一个物种――植生虫草(Phytocordyceps ninchukispora)。
这是我在台湾发现的新属,菌种只有我有,因此它所产生的多醣一定是新的,在新颖性上就没有被挑战的地方。跟我一起完成这项研究的,是我当时收的一位硕士班学生郭俊成(现任嘉南科大教授),他很努力将这些繁复的工作一一完成,建立标准的操作程序,非常感谢他的贡献。
以发酵方式生产「量多质纯」的灵芝多醣
除了分析方法之外,多醣的来源也很重要,如果能在多醣产生过程中控制量多而质纯,後续的分离纯化就会变得很简单。以前在台大农化所学到的发酵方法,很快解决了这些问题。
如前所述,多醣来自真菌细胞壁,可由子实体萃取纯化,但一般子实体所含的代谢物较为复杂,多醣也夹杂後续形成的色素,因此最方便的方式还是来自菌丝体。由於真菌属好氧生物,生长过程需要氧气持续供应,而供氧量会决定液态培养菌丝体的生长速度,所以小样品(500 mL以下)经常采用震荡培养,较大的样品就要采用发酵槽通气培养。
由於制式发酵槽很贵,我们只好自己设计,好用又便宜的克难式发酵槽於是应运而生。也由於这种低搅拌通气方法,不管是植生虫草还是灵芝菌丝,在此系统下产生的菌球就如同珍珠粉圆一般,非常美丽;加上长时间搅拌,细胞壁合成之多醣会释放到发酵液中,最後形成天然活灵芝珍珠粉圆果冻,冷藏後也很可口(详见《健康灵芝》59期第4页)。
这些粉圆果冻先以离心分离菌丝体後,菌液只要经过一个分子筛的设备(这是一款名为Minitan、由Millipore公司制造的中小型超微过滤装置)过滤兼清洗去除小分子,1公升的培养液一天内就可完成;接着再经过冷冻乾燥,每公升约4公克的纯白色粗多醣就出现了。
有了这些原料,後续的研究即可顺利进行,也可免除用酒精沉降法浪费许多酒精的困扰。可惜的是,Minitan装置後来不再生产,以致接下来的研究得另觅他途。
灵芝菌丝体多醣的抗肿瘤研究
在完成原料控制生产及纯化分析後,当然要了解这些多醣到底具有何种功能。检视文献後发现,多醣的生理活性主要在免疫功能之调节,然而免疫学一直是我的弱项,也缺乏研究信心,好在当时的董大成院长对灵芝多醣极有兴趣,我也很乐意成为前段生产纯化分析的生产供应者。
就这样,由灵芝发酵产生菌丝体多醣的免疫抗肿瘤研究又活起来了。这段期间(1995~2000年),以董教授为主要作者在《癌症医学会刊》发表了数篇论文,我受益良多,董教授也成为国内灵芝研究之先驱者。
同时间,我的学长赖敏男教授,对灵芝的液态发酵也深感兴趣,有段时间几乎天天来到我那个很克难又很有趣的实验室,看灵芝珍珠粉圆的生长,和我一起分析残糖含量、菌丝含量、多醣含量变化,并互相讨论放大试验的心得。数年後,赖教授开始建立自己在苗栗双冬的大型发酵厂,成功的生产灵芝菌丝体,我也深感与有荣焉。
重新审视灵芝子实体
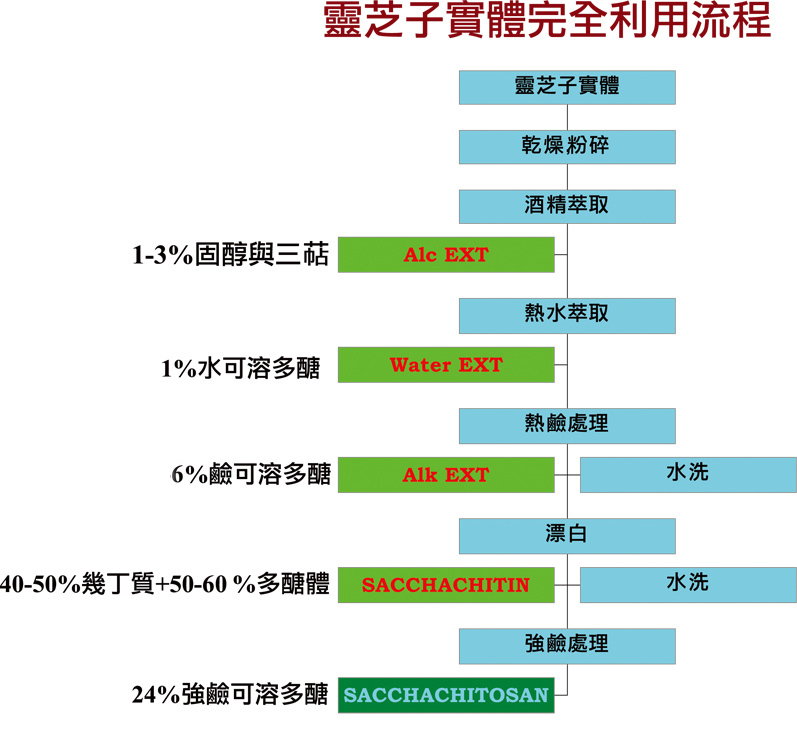
完成这阶段的工作後,我的研究再度回到灵芝子实体。如前所述,灵芝子实体所含的水溶性多醣大概是乾物重的1%左右,相较菌丝体菌液中1公升只能取出4克(0.4%)纯白的多醣,似乎有较高的产率。
然而在操作上,经过乾燥的子实体必须加入两百倍左右的水进行萃取,亦即要取得4克的多醣,得用400克子实体再加80公升的水来萃取,不仅过滤的体积很庞大,所得到的粗多醣成分也相对复杂,常含有子实体的色素,使得多醣呈现咖啡色,这也是一开始我不太喜欢从灵芝子实体萃取多醣的原因。
1990年的某一天,我参加了一个灵芝研讨会,一位从事灵芝栽培的朋友问我:「灵芝子实体无论用水或酒精萃取都会剩下许多残渣(约90%乾物重)。这些残渣除了当有机肥料,有没有其它用途?」这个问题成了启发我回头研究灵芝子实体多醣的灵感。
虽然这段期间我把专注力放在灵芝子实体三类上,但也同样发现,经过水或酒精萃取後的灵芝子实体,在外观上几乎没什麽改变。於是问题又回到真菌细胞壁上,也让我想起指导教授王西华的教诲:「真菌细胞壁就是几丁质和多醣。」
於是我开始思考:「同样都是细胞壁组成,为何不同蕈类有这麽大的差异?」像洋菇、鲍鱼菇、金针菇及香菇都十分滑嫩可口,为何灵芝子实体久咬不烂且难以下咽?这是否是因为几丁质与多醣的比率不同造成的?因为几丁质代表较为坚韧的组织(例如虾蟹、昆虫外壳),多醣则为黏滑吸水性较高的部分。
这些假设很快就得到答案。果然,灵芝子实体含有大量几丁质(40~50%),而洋菇、鲍鱼菇、金针菇等的几丁质含量都小於1%,香菇则为3%。灵芝子实体既然含有这麽多几丁质,为何不假以开发成为生医材料?这於是成了我研究灵芝子实体残渣的开端。
研究子实体残渣的意外收获 ── 硷性可溶多醣
为了得到灵芝子实体几丁质,首先须以热硷(1N NaOH,95℃)破坏蛋白质、核酸、脂质等不需要的杂质,於是就有了热硷萃取液。热硷萃取液本来也是不要的废液,但我们把它回收,以超微过滤分子筛处理,不仅可取出硷可溶多醣,回收率也超过水可溶多醣的六倍左右。
这很可能是灵芝子实体因为高温硷性造成化学键断裂,而使更多的多醣从细胞壁上溶离出来。至於经硷处理的残渣,漂白後就成为白色纸浆状的材质,我们将其命名为SACCHACHITIN,可以做成薄膜,也可以磨成悬浮微粒,是很好的伤口敷料。
基本上,SACCHACHITIN就是几丁质和多醣的复合体。几丁质经过去乙醯化(deacetylation)的过程可以得到几丁聚醣(chitosan),而去乙醯化需要更强的硷(如10 N NaOH)来处理,亦即硷的浓度要比前一次处理强十倍。我们於是把SACCHACHITIN放到强硷中,结果几乎所有的SACCHACHITIN都溶解了。
照理讲,几丁聚醣是硷性的,应该不会溶於强硷中,因此我们推测,SACCHACHITIN除了几丁质及多醣之外,可能还含有相当量的醣酸单体,以致在强硷下可以溶解。
这些假说仍有待进一步证明,不过如果将这些溶解物再以超微过滤回收,更多的多醣就产生了――经过冷冻乾燥後,强硷可溶的多醣高达24%。把灵芝子实体的水可溶多醣(1%)、硷可溶多醣(6%)、强硷可溶多醣(24%)三个处理步骤加起来能产出多达31%的多醣,远超过液态菌丝培养的产值。
(提供/苏庆华)
为了了解这三种多醣是否具有生理活性功能,我们也做了动物试验。结果显示,三种多醣都有抑制S-180肉瘤细胞在肺脏形成结瘤的作用,其中以强硷可溶多醣最明显,可抑制结瘤数目达65%,亦即结瘤数只有对照组的35%。另外再以流式细胞仪分析细胞周期的分布状况,也显示服用强硷可溶多醣的小鼠,正在分裂的肺部细胞(S-180肉瘤细胞)明显下降。
这些结果告诉我们,灵芝实在是好东西,经过不断挖掘,每一个萃取阶段都有好的成分展现出来,而最好的多醣居然留在最後才出现,也完成了对灵芝子实体彻底应用的实例。到目前为止,好像没有厂商对硷性可溶的多醣进行开发,这可能牵涉其中重要的关键设备,需要一套有效率的超微过滤系统。
大型陶瓷超微过滤膜装置技术在产业上之应用已经逐渐成熟,相信不久的将来,这些产品应可达到量产阶段。

