苏庆华教授在本篇文章回顾了他在1986至1990年代初对於三萜类的研究。那段期间,「发现新的三萜类」是灵芝学界的竞逐焦点,没有人力也没有昂贵设备的他,「只好」分析手上各种灵芝标本的三萜类差异,结果发现,每一种灵芝都有独特的三萜类指纹图谱,不会受栽培条件影响,可以作为快速判别灵芝菌种的依据。後来再有人提出类似的主张,已是十几二十年後的事了。
文/苏庆华
◎本文原载於2013年《健康灵芝》第61期 4~6页
1986年九月中,新的学期已经开始,老师和学生一样心情也跟着紧绷,经过漫长的暑假,面对新的学期,要把这学期要上的课大致备好,把暑期中做过的实验数据整理一番,以便在年底向国科会提出新的研究计画。
来北医一年後,大致已将植生虫草多醣的研究做到一个段落,正在思考下一轮的研究主题时,国科会恰巧在筹组一个大型的灵芝研究计画,我很幸运经由董大成教授的推荐加入此一计画,也让我重新回到灵芝的研究领域。
思索我在灵芝研究领域的优势
当时的国际研究环境灵芝已经登堂入室,随着Kohda等学者分离并确定了几个灵芝酸化学构造後,许多日本和荣总萧明熙教授的研究团队,也在接下来几年里陆续从灵芝中发现许多新的三萜类成分。
在这种高度竞争的状况下,能够拥有吃苦耐劳的研究生,成为参与此一竞赛的必要条件,因为研究需要他们从数公斤的材料进行萃取,透过冗长的分离色层管柱分离、纯化、结晶,把这些三萜类成分以高阶的核磁共振仪(MNR)进行分析,再加以判读。
然而,这些条件和设备仪器对一个刚建立小小实验室的我是欠缺的,尤其是动辄数千万的MNR,在当时听起来简直是天文数字(北医在1998年购入高阶MNR,Bruker Avance DRX 500)。因此,在新成分无法与他人竞争的情况下,回头反思自己到底有什麽优势,而当时我所拥有的就是一些灵芝菌种,以及从各地采集的子实体标本。
这些标本有些很容易辨识,大多数则雷同或小有差异,亦即以子实体形态作为分类依据其实是很困难的。我们也采集担孢子,利用光学显微镜及扫描式电子显微镜,试图区分这些标本的归属,但除了紫芝群具有苦瓜型的担孢子外,老实说,连我也无法区别出其它种类。因此,在台湾大学许瑞祥教授的DNA指纹技术尚未建立之前,化学方法或许可作为一项参考。
分析灵芝成分的初步成果,以及衍生的七个问题
一开始我就用最简单而快速的方法,针对这些标本进行薄层色层分析(TLC)。TLC可以在很短的时间内得知重要的定性资讯,只要大约2 g样品,以100 ml甲醇或其它溶剂萃取後浓缩至1.5 ml,再取2 μl 点在薄层分析片上,以甲醇:氯仿:水=30:4:1的体积比,展开、烘乾、呈色、照相就可以了。我那时随便取了几个样品进行初步分析,结果十分令我惊讶【图1】。
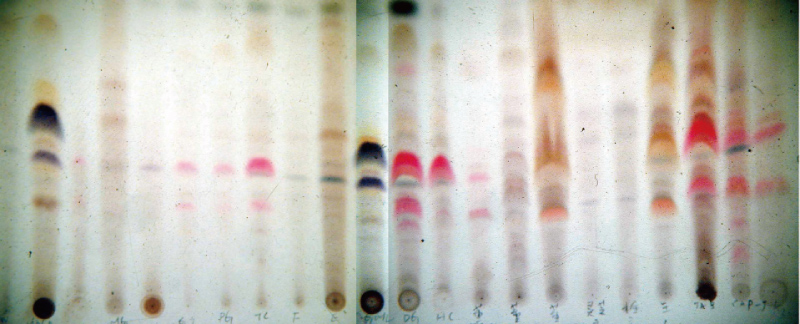
图1 灵芝子实体逢机取样之TLC(薄层色层分析)图谱。(摄影/苏庆华)
依据我原先的预期,灵芝之间的成分差异应该是少许的,或者说是渐进的改变,没想到其间差异是那麽大。这样的结果也让我开始想到一连串的问题,其中最主要的一些辩证包括:
问题1: 灵芝子实体的三萜化学图谱是否有固定的指纹,还是会因为环境而改变?
问题2:三萜化学图谱如有固定指纹,是否会因种或品种而有所差异?
问题3:是否因为灵芝种之间的成分差异,而影响灵芝的功效?
问题4:三萜化学指纹是否可作为分类之依据?
问题5:不同种的灵芝能否交配,交配後下一代的指纹又是如何?
问题6:所有发表之灵芝三萜全来自同一个种?
问题7:能否透过育种得到大量而单一的三萜?
灵芝子实体的三萜化学图谱有固定指纹
以上这些问题一直围绕在後续研究中,也主导我对灵芝的研究方向与看法。针对第一个提问,大概花了三年的时间把同一个野生种,种在不同的地方,包括南投、嘉义,还有台北我的实验室中,而我的实验室也用木屑、糙米等不同的培养基,进行第二代及第三代的灵芝栽培。结果所呈现出来的图谱是完全一样【图2】,再加上後来使用高效液态色呈分析(HPLC)图谱,也同时解答三萜含量的多寡会因采收期而改变。
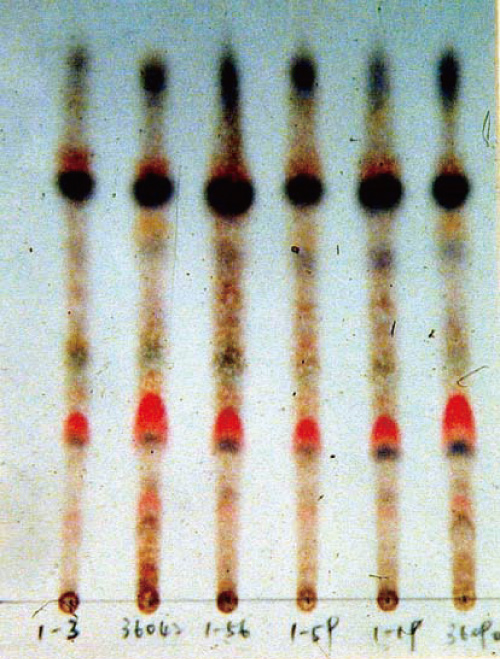
图2 以松杉灵芝为例,在不同培养基、种植地点、不同世代栽培之子实体,
呈现相同的TLC三萜指纹图谱。(摄影/苏庆华)
当初我把想法提出来和大家讨论时,也有不同的意见,传统上认为三萜属於二级代谢产(secondary metabolites),会因为培养环境而改变,但几年後大家也逐渐认同我的看法。
随後我逐渐有了研究生,其中詹美华同学花了很多时间把灵芝酸A、B、C2、D都纯化出来,有了纯化的标准品分析就方便多了。此外,美华同学也证明灵芝酸C2为灵芝保肝功能的主要成分。
杨依珍同学则分析了三百多个样品,并将之归纳成十八种类型【图3】。有趣的是,这十八种类型大致可区分为两大类,图3右侧的八种(K-R)都具有两个红色呈色,分别为灵芝酸B(较上方)及C2(较下方);左侧十种则不呈现此二成分,显示灵芝子实体形成过程中,有稳定的三萜形成模式。
在当时以DNA指纹监定技术尚未成熟前,三萜类指纹也不失为一种判别灵芝菌种的方法,更何况灵芝以健康食品作为标的,能够呈现灵芝产品的主要成分及来源也是很有意义的。
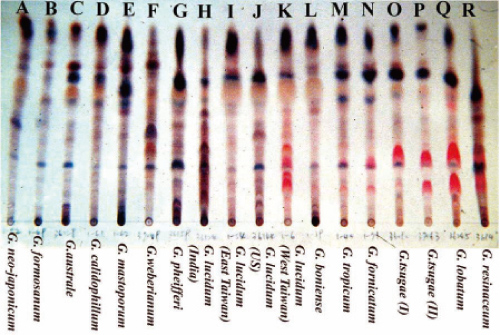
图3 三百多个灵芝子实体以TLC图谱分成十八种类型。(摄影/杨依珍)
灵芝三萜的保肝与抗血癌作用
杨依珍同学除了很仔细地把我手上拥有的子实体标本进行整体分析外,还把这十八种灵芝的萃取物,与肝脏保护功能和抗血癌细胞联结在一起,显示肝脏保护在所有灵芝种类多少都有具备,并无太显着的差异【图4】,但对於血癌细胞的抑制,则有极明显的不同,其中以来自西欧的两个种,Ganoderma recinaceum和G. pheifferi,对血癌细胞的抑制能力特别明显【图5】。
我们也针对这两个种的灵芝在实验室中进行尝试性的栽培,G. pheifferi生长情况很不好,只采得极少数的子实体,而G. recinaceum则生长茂盛,与一般的灵芝栽培种无异。到了2000年,谢翊翎同学证实了血癌细胞的抑制成分为某一个三萜类。这三位同学的努力也回答了我先前提出的第2、3、4个问题,答案都是肯定的。
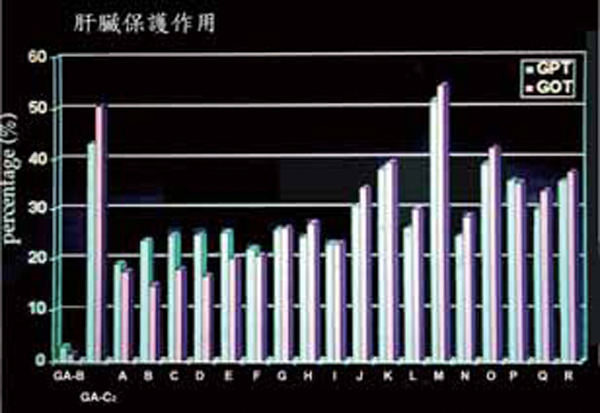
图4 十八种类型灵芝子实体对小鼠四氯化碳造成的肝功能障碍之复原百分比。
(资料来源/杨依珍,台北医学院天然物医学研究所硕士论文,1994)
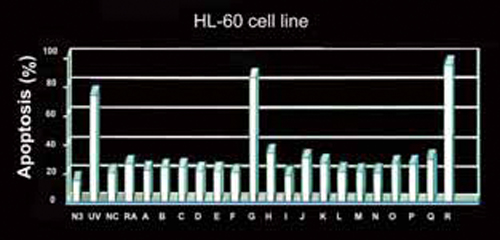
图5 十八种类型灵芝对HL-60血癌细胞株致凋亡之百分比。
(资料来源/杨依珍,台北医学院天然物医学研究所硕士论文,1994)
待解习题:不同种交配後代的三萜指纹?
至於提问5,我们也有机会进行一些尝试。要把灵芝进行交配,首先要得到不同种(或品种)的单核菌丝菌种,但由於灵芝的有性孢子发芽率很低,而且有些甚至根本不发芽,例如G. tsugae(图3 O、P),所以在交配上有一定的困难度。我们很幸运能在几个子实体,例如G. lucidum(图3 H、I、J、K)释放出来的孢子分得多数的单核菌株,不同菌株之间也可以交配形成双核菌株。
这些双核菌株经过栽培也形成子实体,初步分析这些子实体也和亲本不相同,但由於子实体的数量有限,无法得到足够的三萜萃取物,研究很可惜没能继续。如有机会,以现在的成分分离、纯化、监定技术与经验,一定有机会解开这个谜题,尤其配合DNA基因体分析,会让此研究变得很有趣。
目前发表的三萜成分,应来自多种灵芝
第6个提问到目前为止也没有明确的答案,但我相信是否定的。因为发表这些新成分的都是化学家,并没有真菌学家参与,因此他们很可能以一些野生标本或市售的商品作为研究材料,论文所冠的学名虽然千篇一律都是G. lucidum,但是从单一G. lucidum所呈现的指纹,应该没有已发表的成分那麽多,因此推测这些成分应是类似种类的大集合,但即使如此,也不会因而影响灵芝拥有许多保健功能所建立的美誉。
第7个提问与第5个则息息相关,三萜的育种是未来重要的课题,包括具有功能性三萜类生物合成途径的了解,以及基因和基因体学的探讨,这也是灵芝研究真菌学家未来努力的方向。
延伸阅读
1. 〔苏庆华谈三萜〕灵芝酸B及C2在各种灵芝中之含量
2. 〔2015灵芝研发与应用学术研讨会〕陈若芸演讲:
灵芝属真菌化学成分和质量控制方法研究
3. 更多关於苏庆华的文章

